METABOLISMO
CELULAR: RESPIRACIÓN Y FOTOSÍNTESIS
1. GENERALIDADES
El metabolismo
es el conjunto de transformaciones e intercambios que ocurren en el interior
celular, cuyos objetivos son: obtener energía utilizable (ATP) para fabricar
sus componentes celulares y realizar sus funciones vitales.
Como ya sabemos, el metabolismo celular comprende
dos tipos de procesos: el catabolismo (procesos degradativos) y el anabolismo
(procesos constructivos). Si comparamos ambos:
CATABOLISMO
|
ANABOLISMO
|
·
Fase degradativa: moléculas orgánicas
complejas y reducidas (glúcidos, lípidos y proteínas) son degradados a
compuestos finales más sencillos y oxidados.
·
Son procesos exergónicos: liberan energía libre
utilizada para sintetizar ATP (ADP + Pi).
·
Implican oxidación de moléculas orgánicas que
liberan electrones y átomos de H ricos en energía, que se almacenan como
coenzimas reducidos (NADPH, NADH, FADH2).
·
Son rutas convergentes: a partir de muchos
compuestos iniciales se forman unos pocos productos finales.
|
·
Fase constructora (o de biosíntesis):
moléculas sencillas y oxidadas son utilizadas para construir moléculas
complejas y reducidas.
·
Son procesos endergónicos: requieren energía
procedente de la hidrólisis del ATP a ADP + Pi.
·
Implican reducción de distintas moléculas a
partir de electrones y protones cedidos por coenzimas previamente reducidos
(NADPH, NADH y FADH2).
·
Son rutas divergentes: a partir de unos pocos
precursores se forma una gran variedad de productos finales.
|
Los procesos catabólicos y anabólicos no
ocurren ni de forma simultánea, ni en el mismo lugar de la célula, pero están
acoplados.
El estudio del metabolismo se aborda en forma de rutas metabólicas, que son secuencias
de reacciones químicas que relacionan dos metabolitos importantes. Ya sabemos
que, según el origen de la materia y la energía, se distinguen células:
autótrofas, heterótrofas, fotosintéticas y quimiosintéticas. Son los organismos
Fotolitotrofos que utilizan la luz como fuente de energía y el CO2 como fuente
de materia y lo constituyen las algas cianoficeas y los vegetales superiores;
Fotoorganotrofos (luz y materia orgánica)
formado por las bacterias purpúreas no sulfuradas; Quimiolitotrofos
(reacciones redox y CO2) bacterias desnitrificantes y Quimiorganotrofos (reacciones
redox y materia orgánica) y lo constituyen animales y hongos.
Transformaciones energéticas
La mayoría de las reacciones
metabólicas llevan aparejados intercambios de energía. En los seres vivos los
intercambios de energía solo se pueden hacer en forma química. Intercambiando
enlaces químicos entre sustratos, la energía nunca puede quedar libre pues
provocaría aumentos de temperatura en las células, lo que desnaturalizaría a
las proteínas…. La energía se intercambia mediante enlaces de alta energía con
el ión fosfato (PO43- = Pi) en el
seno de un nucleótido, el AMP generalmente, al que para introducirle un segundo
Pi hay que romper la repulsión eléctrica de la molécula y formar el ADP y aún
más para formar el ATP ……. La misma cantidad de energía que se libera al
romperse, transfiriéndose esta energía a otros enlaces químicos recién
formados.
En los
seres vivos las reacciones que necesitan o que liberan energía son reacciones
de oxido-reducción (una molécula
está reducida u oxidada según sus cantidades de O, H, electrones o energía). Los
electrones y el H son sinónimos de energía y como ella tampoco pueden quedar
libres pues reducirían indeseablemente a
la materia orgánica por lo que cuando se intercambian, nunca quedan sueltos,
son aceptados por los coenzimas redox que son sustancias que actúan como
transportadores tomando 2H del sustrato, 2 protones y dos electrones (NAD,
NADP, FAD, FMN ….) Son moléculas muy reducidas que solo aceptan electrones de
alta energía (con alto potencial redox), capaces de reducir, incluso a
sustancias muy reducidas como estas coenzimas. Es el poder reductor.
En las reacciones redox, los electrones pasan de
un átomo o molécula a otro u otra. En general, una oxidación consiste en
la pérdida de electrones, mientras que una reducción consiste en su ganancia.
Naturalmente, para que un compuesto pueda oxidarse tiene que haber otro que se
reduzca, por lo que estas reacciones siempre están acopladas, de ahí que
reciban el nombre de reacciones de oxido-reducción.
Modalidades de fosforilación
La fosforilación es la adición de un
grupo fosfato inorgánico (PO43- = Pi) a
cualquier otra molécula. En el metabolismo, este es el mecanismo básico de
transporte de energía desde los lugares donde se produce hasta los lugares
donde se necesita.
La fosforilación más importante es
la fosforilación de ADP: adición de
un grupo Pi a un ADP para formar ATP (ADP + Pi → ATP + H2O), actuando este ATP como la “moneda
de cambio energético” del metabolismo.
Existen distintas modalidades de
fosforilación de ADP:
·
FOSFORILACIÓN A NIVEL DE SUSTRATO: transferencia de un Pi desde un compuesto
fosforilado a un ADP. Para hacerlo, utiliza la energía liberada tras la
hidrólisis del grupo fosfato. En la glucolisis, el ácido fosfoenolpirúvico,
cede su Pi directamente al ADP para formar ATP quedando como ácido Pirúvico.
·
FOSFORILACIÓN
ASOCIADA A UN GRADIENTE QUIMIOSMÓTICO: se acopla al transporte de
electrones (de alta energía, cedidos por la molécula que se oxida) a
través de una “cadena transportadora de electrones”, en cuyo transcurso van
perdiendo energía, que se utiliza para impulsar protones a través de una
membrana, generando un gradiente electroquímico de protones a ambos lados de la
membrana. Este gradiente permite que los H+ pasen luego a través de
una ATP-sintetasa, que utiliza la energía liberada por el transporte a favor de
gradiente de H+ para sintetizar ATP a partir de ADP y Pi. A su vez,
existen dos tipos:
o
Fosforilación oxidativa: ocurre en la mitocondria, a nivel de la
membrana mitocondrial interna.
o
Fotofosforilación: ocurre en los cloroplastos, a nivel de la
membrana tilacoidal y durante la fase luminosa de la fotosíntesis.
2. CATABOLISMO
Como ya hemos comentado anteriormente, en el
catabolismo las moléculas orgánicas complejas se van degradando en otras más
sencillas, lo que conlleva una liberación de energía que permite la formación
de ATP.
En general, el catabolismo lo constituyen procesos
de óxido-reducción en los que intervienen fundamentalmente enzimas
deshidrogenasas. Así pues, la oxidación de moléculas orgánicas va liberando
electrones y, dependiendo de la naturaleza del aceptor final de electrones,
se diferencian dos modalidades de
catabolismo:
·
FERMENTACIÓN: es
una oxidación incompleta, en la que el aceptor final de electrones es un
compuesto orgánico. Es, pues, un proceso anaerobio (ya que no interviene
el oxígeno).
·
RESPIRACIÓN CELULAR: es una oxidación completa de compuestos
orgánicos, en la que el aceptor final de electrones es una sustancia
inorgánica. Según la naturaleza de este aceptor inorgánico se distinguen:
o
Respiración aerobia: el aceptor final es el oxígeno (que al reducirse
forma agua).
o
Respiración anaerobia: el aceptor final es una sustancia diferente al
oxígeno (ej: NO3-, SO42- o CO2).
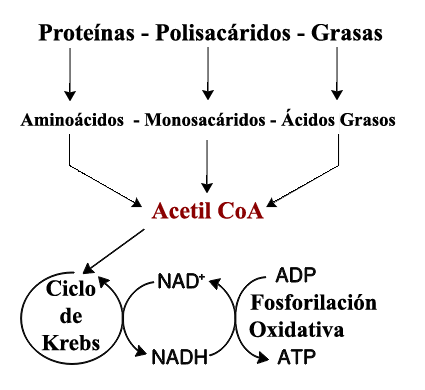
Esquema general del catabolismo:
Como puede observarse en el dibujo,
los alimentos que se oxidan pueden tener distintos orígenes,
básicamente glúcidos, lípidos y proteínas; pero, independientemente del origen,
todas las rutas catabólicas convergen en un intermediario metabólico (el
Acetil Co-A), el cual completa su oxidación hasta CO2 en el Ciclo de
Krebs.
Los electrones liberados como
consecuencia de las sucesivas oxidaciones son cedidos a una “cadena transportadora
de electrones”, que impulsa finalmente la formación de ATP por fosforilación
oxidativa.
2.1 CATABOLISMO
DE GLÚCIDOS.
Las rutas metabólicas por la que la célula degrada
los glúcidos para la obtención de energía son la respiración celular aerobia
(mayoritaria), respiración anaerobia y la fermentación; en todas
ellas, la degradación comienza con un proceso común: la Glucolisis.
GLUCOLISIS
Es una ruta prácticamente universal, que ocurre
en el citosol, consistente en una secuencia de 10 reacciones metabólicas,
en las que, a partir de una molécula de Glu, se obtienen dos de piruvato y 2
ATP (mediante fosforilaciones a nivel de sustrato).
La glucolisis se puede dividir en
tres etapas:
·
Etapa de Fosforilación: la molécula de Glu se activa por la unión de 2 Pi
(procedentes de la hidrólisis de 2 ATP), y se isomeriza dando lugar a la Fru-1,6- Difosfato, que se
escinde en 2 gliceraldehído-3-P.
·
Etapa de Oxidación: cada molécula de gliceraldehído-3-P se oxida
dando una molécula de 1,3-difosfoglicerato, con la incorporación de un fosfato
inorgánico. En esta oxidación se obtiene poder reductor, en forma de NADH.
·
Etapa de Síntesis de ATP: ocurre la primera síntesis de ATP, mediante a la
transferencia de uno de los grupos Pi del 1,3-difosfoglicerato al
ADP; como consecuencia, se libera una molécula de 3-fosfoglicerato, que se
transforma en piruvato mediante la liberación de su Pi a un nuevo
ADP (lo que produce un nuevo ATP).
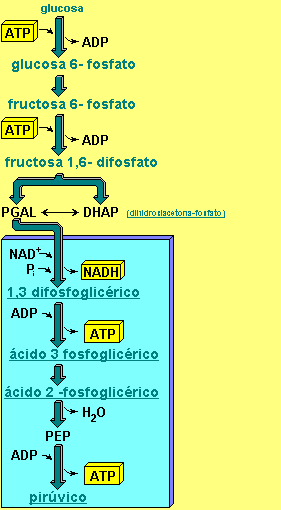
Observando el diagrama de la glucolisis, se puede
deducir el balance global de la ruta:
Glu + 2 ADP + 2 Pi + 2 NAD+ → 2 piruvato
+ 2 ATP + 2 NADH + 2 H+ +
2 H2O
Podemos concluir que la eficacia de la glucolisis como ruta energética es
muy baja, puesto que únicamente tiene un rendimiento neto de 2ATP/Glu. Pero además, genera poder
reductor (2 NADH), el cual, en caso
de que la célula continúe el proceso de respiración, originará más ATP.
RESPIRACIÓN AERÓBICA
Es la oxidación
total del producto final de la
glucolisis (piruvato), lo cual va a liberar una gran cantidad de electrones
que van a ser finalmente cedidos al O2. Para facilitar su estudio, se divide en tres
etapas:
·
Formación del Acetil-CoA:
El piruvato obtenido en la glucolisis es conducido
desde el citoplasma a la matriz mitocondrial, unido a transportadores
específicos que le permiten atravesar la doble membrana mitocondrial, donde se
decarboxila oxidativamente obteniéndose Acetil-CoA y 1 NADH. Es una secuencia
de reacciones catalizadas por el complejo multienzimático de la Piruvato deshidrogenasa
que deshidrogena y descarboxila al Pirúvico y combina el ácido acético
resultante a su transportador, la coenzima A, formándose acetil Co A
(combustible de la respiración celular) y un NADH.
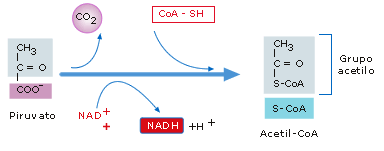
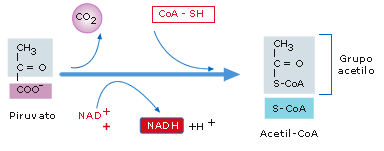
·
Ciclo de Krebs: también
llamado “Ciclo de los ácidos tricarboxílicos”,
es un conjunto cíclico de reacciones de descarboxilaciones y
deshidrogenaciones en las que se produce la oxidación completa de Acetil-CoA
hasta CO2 y 8 H (ocurre en la matriz mitocondrial). Los
electrones que se liberan en la oxidación son captados por las coenzimas FAD y
NAD+ liberándose, por cada vuelta de ciclo: energía (1 GTP convertible a ATP), poder
reductor (1 FADH2, 3 NADH) y 2 CO2.
El Ciclo de Krebs es el núcleo del
metabolismo intermediario, tratándose de una vía anfibólica, es decir, resulta clave tanto para procesos
catabólicos como anabólicos. Consiste basicamente, en descarboxilaciones y
deshidrogenaciones del ácido acético.

·
Fosforilación oxidativa: tiene lugar en la membrana mitocondrial interna
e incluye tres etapas:
o
Transporte electrónico:
En este proceso, los coenzimas reducidos (NADH y
FADH2) ceden sus electrones a una “cadena transportadora de
electrones”, donde “caen” a favor de gradiente de potenciales de
óxido-reducción hasta el aceptor final (O2).
La cadena transportadora está formada por un
conjunto de moléculas capaces de reducirse y oxidarse, dispuestas según un
gradiente de potenciales de óxido-reducción, organizadas en 4 complejos
(insertos en la membrana mitocondrial interna):
 §
Complejo I (NADH-deshidrogenasa mitocondrial), que acepta protones
y electrones del NADH y los cede al…
§
Complejo I (NADH-deshidrogenasa mitocondrial), que acepta protones
y electrones del NADH y los cede al…
§
Complejo II (Ubiquinona): que acepta también electrones y
protones del FADH2 y los cede al…
§
Complejo III (citocromo b-c1. Los citocromos poseen
un grupo hemo por lo que solo acepta electrones, pasando su Fe férrico a ferroso,
quedando los protones en la matriz) que cede los electrones al…
§
Complejo IV (citocromo-oxidasa): que cede los electrones al O2
, que es el aceptor último de los electrones quedando como un anión
superoxidado O2- que con 2 H+ se reduce formando H2O.
o
Formación del gradiente quimiosmótico: a disminución del potencial redox de los
electrones, la energía liberada por el transporte electrónico se utiliza para bombear
protones (H+), translocar protones, a nivel de los complejos I,
III y IV hacia el espacio intermembrana, creando un gradiente
electroquímico de protones entre el espacio intermembrana y la matriz
mitocondrial (NADH/2 e-/ translocación de 6 H
+ FADH2/2 e-/ translocación de 4 H+)
o
Síntesis de ATP: dada la situación de inestabilidad que genera el
gradiente electroquímico de protones, los protones van a regresar a la
matriz atravesando el canal interno de una proteína transmembrana (ATP
sintetasa); ello disipa el gradiente de protones y permite utilizar la
energía liberada (por el transporte a favor de gradiente) para la fosforilación
del ADP y, por tanto, producción de ATP (fosforilación oxidativa). Los
H+ entran por la base hidrófoba, atraviesan el pedúnculo Fo y salen por la esfera catalítica F1 provocando un cambio en la estructura de la
proteína que le permite fosforilar el ADP con un Pi y a los H+ volver a la
matriz. Un símil podría ser, el de un generador eléctrico a vapor en el
que el calor que calienta el agua
hasta vapor sería el flujo de los
electrones que produce el gradiente
electroquímico (vapor) cuya
presión hace girar una turbina (ATP
sintetasa) que es la produce la electricidad(ATP)
Rendimiento energético de la Respiración Aerobia
La
Respiración Aerobia es
un proceso energéticamente muy eficiente,
ya que: se produce oxidación completa de los C hasta CO2 y porque
la diferencia entre los potenciales de óxido-reducción del sustrato
oxidable y el aceptor final de electrones es muy grande.
Para poder calcular el balance
energético de la respiración aerobia, es necesario conocer que, en la
fosforilación oxidativa, cada par de electrones cedidos desde el NADH origina 3
ATP mientras que los cedidos por el FADH2 se originan sólo 2 ATP.
Así pues, a partir de cada molécula de Acetil-CoA se forman 12 ATP, por lo que por cada molécula de
glucosa que inició la glucólisis se obtienen 24 ATP.
Rendimiento energético de la: OXIDACIÓN COMPLETA
DE LA GLUCOSA
La producción neta de ATP en la oxidación completa de la glucosa
(Glucólisis + Respiración aerobia) es de
36-38 ATP, dependiendo de la
“lanzadera” que se utilizase para incorporar los electrones contenidos en
los NADH generados en el citosol (durante la glucolisis):
·
Lanzadera Glicerol-fosfato: el NADH citoplasmático cede sus electrones a
FAD mitocondrial, con lo que la cadena respiratoria sólo rendirá 2 ATP. El
balance neto en este caso sería de 36 ATP.
·
Lanzadera Malato-Aspartato: el NADH citosólico cede sus electrones al NAD+
mitocondrial, por lo que el balance neto sería de 38 ATP.

RESPIRACIÓN
ANAERÓBICA
Mientras
que la respiración aeróbica de moléculas orgánicas requiere la presencia de O2
y es típica de organismos superiores, existe otra modalidad de respiración,
la anaeróbica, típica de procariotas
(Bacterias) que no requiere la presencia de O2.
Se
trata, por tanto, de una ruta prácticamente igual a la respiración aeróbica
(con cadena transportadora de electrones donde pueden regenerar el NAD+)
pero con la diferencia de que, en lugar de utilizar O2 como aceptor final de electrones,
utiliza NO3-,
algún compuesto orgánico o el hierro. Al igual que en la respiración
aerobia, existe una oxidación completa del sustrato y se sintetiza ATP
mediante fosforilación oxidativa.
FERMENTACIÓN
Es
una ruta metabólica que ocurre en el citoplasma, mediante la que las células
obtienen energía en condiciones
anaeróbicas (el aceptor final de electrones es un compuesto orgánico y no
el O2), por oxidación parcial
de compuestos orgánicos (sobre todo glúcidos); la síntesis de ATP se produce
por fosforilación a nivel de sustrato.
La
fermentación surge, a lo largo de la evolución, como una estrategia que
permitió a los organismos vivir en una atmósfera carente de oxígeno. Actualmente,
es una ruta metabólica utilizada por:
ü
Microorganismos
(básicamente bacterias) anaerobios estrictos o anaerobios
facultativos.
ü
Algunas células
eucariotas en condiciones de anaerobiosis (ej: células musculares).
Como
ya se ha comentado, los sustratos a fermentar son normalmente glúcidos y el
proceso tiene lugar en dos etapas:
·
ETAPA DE OXIDACIÓN (Glucolisis): oxidación de la glucosa hasta piruvato. Se
consume 2 ADP y 2 NAD+ y se produce 2 ATP y 2 NADH.
·
ETAPA DE REDUCCIÓN: reducción del piruvato obtenido en la anterior
etapa mediante la oxidación de los 2 NADH obtenidos también anteriormente. Ello
provoca la regeneración del NAD+ (lo que permite que la glucólisis
no cese). Dependiendo de cuál sea el producto final, existen dos tipos de
fermentación:
o
Fermentación Láctica: el piruvato se reduce hasta ácido láctico, al
aceptar
los
electrones del NADH producido en la glucólisis; es una reacción catalizada por
la enzima “Lactato Deshidrogenasa”.
El balance global de la reacción es:

Glu + 2 ADP + 2Pi → 2 lactato + 2 ATP
Este tipo de fermentación la realizan las bacterias lácticas, como las del género
Lactobacillus y Streptococcus, que provocan el
agriado de la leche y son las responsables de la obtención industrial de sus
derivados: queso, yogur, etc. También la realizan las células musculares durante un ejercicio intenso o prolongado en los
que el aporte de oxígeno es insuficiente como para realizar respiración
aeróbica; en este caso, los pequeños cristales de lactato en las fibras
musculares provocan las conocidas agujetas.
o
Fermentación Alcohólica: el piruvato sufre una doble reacción; en primer
lugar se descarboxila
dando acetaldehído; en segundo lugar, el acetaldehído se reduce hasta etanol
(en una reacción catalizada por la “Alcohol
Deshidrogenasa”), al aceptar los electrones del NADH producido en la
glucolisis.
El balance global de la reacción es:

Glu + 2 ADP + 2 Pi → 2 etanol
+ 2 CO2 + 2 ATP
Este
tipo de fermentación la realizan principalmente levaduras, entre ellas la más conocida “Saccharomyces cerevisiae”, utilizada tanto para la producción
del vino, el pan, la sidra y cerveza (en el caso del pan, el CO2 y
el etanol se eliminan en el proceso de cocción).
Otras fermentaciones, la heteroláctica mixta (láctico,
etanol y CO2)
y otras que finalizan en metano, acetona, succínico, propiónico, butírico…… lo
que permite comprender la importancia de las bacterias en la biotecnología y
bioenergética, solo hay que “echarles de comer” glúcidos naturales no
comestibles para obtener productos de interés comercial o medicinal.
Rendimiento
energético de la
Fermentación
Como
se puede ver en el balance global tanto de la fermentación láctica como de la
alcohólica, el rendimiento energético
es bastante bajo: sólo 2 ATP en comparación con los 36-38 ATP
obtenidos mediante la respiración celular. Ello es debido a que, al tratarse de
una oxidación incompleta, los productos finales son moléculas orgánicas que
conservan aún mucha energía.
Como
también se puede observar, en las fermentaciones no existe producción neta de poder reductor (NADH) ya que el que se
produce durante la fase de oxidación se utiliza para reducir al producto de la
glucólisis en la fase de reducción.
2.2 CATABOLISMO DE
LÍPIDOS.
Como
ya sabes, los lípidos (más concretamente los triglicéridos) son la principal reserva energética en animales,
acumulándose especialmente en el citosol de las células del tejido adiposo. Las
dos razones principales son:
ü
Almacenan más
energía que el Glucógeno (9 Kcal/g frente
a las 4 Kcal/g que aportan los glúcidos).
ü
Al ser hidrofóbicos, no
se almacenan hidratados (a diferencia del Glucógeno que es hidrofílico) por
lo que pesan menos. Aspecto importante para facilitar la movilidad animal
Cuando la célula necesita un aporte
energético mayor del habitual, o no dispone de glúcidos, degrada las grasas. El catabolismo de los triglicéridos comienza por su hidrólisis, realizada
por enzimas lipasas presentes
en el intestino delgado, obteniéndose
glicerina y ácidos grasos. La glicerina se convierte fácilmente en
gliceraldehido-3-P y continúa la ruta de la glucolisis. Los ácidos grasos siguen una ruta especial, llamada β-oxidación.
Esta
oxidación ocurre en la matriz mitocondrial de todas las células (excepto en las
del cerebro y el riñón). El paso de los ácidos grasos desde el citoplasma a la
matriz mitocondrial requiere una previa activación del ácido graso, mediante la
unión de un CoA (requiere energía aportada por la hidrólisis de 1 ATP→AMP + Pi) originando
Acil-CoA. Es entonces cuando el ácido graso activado (Acil-CoA) atraviesa la
doble membrana mitocondrial, mediante un sistema de transporte mediado por una
molécula transportadora, llamada carnitina.
Ya
en el interior de la matriz mitocondrial, comienza la β-oxidación del Acil-CoA,
mediante una serie de cuatro reacciones sucesivas.
En
cada ciclo de β-oxidación se desprende una molécula de Acetil-CoA, 1 FADH2
y 1 NADH, quedando el ácido graso inicial con dos carbonos menos. Este se
activa de nuevo e inicia una nueva β-oxidación, repitiéndose el proceso hasta
que se oxida totalmente el ácido graso. Por esto los ácidos grasos naturales
tienen un número par de átomos de carbono.
Finalmente,
las moléculas de Acetil-CoA se incorporan al Ciclo de Krebs, donde son oxidadas
completamente, y todas las moléculas de NADH y FADH2 ingresan en la
cadena transportadora respiratoria, produciendo una gran cantidad de ATP por fosforilación
oxidativa.
Rendimiento energético de los lípidos
La
oxidación completa de los triglicéridos libera una gran cantidad de energía debido a que su cadena hidrocarbonada está
muy reducida. Por ejemplo: la triestearina, que se descompone en glicerol
y 3 ácido esteárico (18 C),
origina 460 ATP.
NOTA: cuando el
nivel de glucosa en sangre es bajo,
el hígado puede transformar el acetil-CoA de la β-oxidación en unos compuestos
químicos especiales (cuerpos cetónicos),
que pueden atravesar las membranas celulares y utilizarse como combustible de
muchos órganos; este proceso es de vital
importancia para determinados órganos vitales, tales como el cerebro, el
corazón o el riñón, que normalmente sólo utilizan glucosa.
3. ANABOLISMO
Como ya sabemos, el anabolismo es el conjunto de
procesos constructivos que suceden en la célula en los que se produce la
síntesis de moléculas complejas a partir de otras sencillas. Estos procesos
endergónicos necesitan la incorporación de energía.
Para sintetizar moléculas biológicas,
y a diferencia que en el catabolismo, la célula necesitar llevar a cabo una
serie de reacciones de reducción que
irán acopladas a la oxidación del NADH o del NADPH.
Se pueden diferenciar dos grupos de procesos anabólicos:
·
Los que permiten fabricar
moléculas orgánicas a partir de
inorgánicas; son exclusivos de organismos autótrofos y, en función de cuál
es la fuente primaria de energía, se diferencian:
o
FOTOSÍNTESIS:
utilizan la energía de la luz (ej: plantas, algas y cianobacterias).
o
QUIMIOSÍNTESIS: utilizan la energía liberada en las reacciones
químicas exergónicas (ej: bacterias del nitrógeno, del azufre o las del
hierro).
·
Los que permiten
fabricar moléculas orgánicas complejas a
partir de moléculas orgánicas sencillas:
o
Anabolismo de
Glúcidos: GLUCONEOGÉNESIS y GLUCOGENOGÉNESIS.
o
Anabolismo de
Lípidos.
o
Anabolismo de
Proteínas.
3.1. FOTOSÍNTESIS
La fotosíntesis es un proceso de
nutrición autótrofa por el que se forma materia orgánica (monómeros de los
polímeros orgánicos) por reducción de materia inorgánica (CO2,
H2O y sales minerales), utilizando la energía luminosa. Se trata
pues de un proceso anabólico utilizado tanto por plantas, algas, como algunas
bacterias.
La vida en el planeta depende de los organismos fotosintéticos ya que
son los únicos capaces de fabricar la materia orgánica de la que
dependen el resto de los organismos de las cadenas tróficas (Consumidores y
Descomponedores). Por lo que la importancia biológica de la fotosíntesis
es fundamental para el mantenimiento de la vida sobre la tierra y la evolución
de las especies.
Dado que se trata de una reacción redox, requiere
un dador de electrones y un aceptor. En función de qué moléculas actúen como
dadores y aceptores de electrones, se diferencian dos tipos de fotosíntesis:
·
Oxigénica (o vegetal):
el dador es el H2O y el aceptor el CO2. La
fotólisis del agua es la responsable del desprendimiento
de oxígeno molecular (O2) a la atmósfera, que resulta vital para
la vida de los organismos que presentan metabolismo aerobio. Es realizada por
plantas, algas y cianobacterias. Fotolitotrofos.
·
Anoxigénica (o bacteriana): el dador es el sulfuro de hidrógeno o el acido
láctico y el aceptor es el nitrato o el sulfato. Puesto que no
utiliza el agua como dador de electrones no se desprende O2 a la atmósfera.
Es realizada por determinadas bacterias. Quimiolitotrofos y Fotoorganotrofos.
FOTOSÍNTESIS OXIGÉNICA
Tiene lugar en los cloroplastos y se
lleva a cabo en dos etapas:
·
FASE LUMINOSA: se denomina así porque ocurre en presencia de
luz, en la membrana tilacoidal, y en ella unas moléculas
fotorreceptoras (pigmentos fotosintéticos) captan la energía de la luz y la
transforman en energía química (ATP
y NADPH).
Los pigmentos
fotosintéticos constituyen el 12% de la composición de la membrana
tilacoidal. Son lípidos isoprenoides, carotenos, xantofilas y fitol (forma parte de la clorofila,
heteroproteina con grupo hemo como el de la Hemoglobina pero con
Mg en lugar de Fe). Cada pigmento absorbe la luz de una determinada longitud de
onda gracias a sus muchos dobles enlaces, electrones deslocalizados capaces de
absorber energía y alcanzar un orbital electrónico mas energético y después
desprenderla volviendo a su orbital (La clorofila , in vitro, emite
fluorescencia tras ser excitada con un fotón)
La luz son ondas, radiaciones electromagnéticas emitidas
por el Sol, por ejemplo, y que se trasladan como una onda, con un movimiento
ondulatorio y que cuando interacciona con la materia (absorción o emisión por
átomos o moléculas) se comporta como si estuviese formada por paquetes de
energía, cuantos o fotones (se comporta como los electrones o protones aunque
sin masa ni carga como ellos) tiene un
doble carácter de onda y de partícula. Como todas las ondas, sonoras, sísmicas
….. comparte sus propiedades, dirección, frecuencia, longitud de onda, intensidad (energía) …… Precisamente a mayor
longitud de onda menor frecuencia y energía (luz infrarroja, ondas de radio
microondas) y viceversa (rayos gamma, X, ultravioletas) …… demasiado
energéticas y las otras demasiado poco para los seres vivos pero las
intermedias, (la luz visible, del
rojo al violeta, de 400 a
700 nanometros son perfectas para la
fotosíntesis)
Tiene lugar en tres procesos:
o
Captación de energía
luminosa:
Lo lleva a cabo el denominado complejo antena, que es un conjunto formado por proteínas y pigmentos
(clorofilas a y b; y carotenoides:
β-caroteno y xantofilas, que absorben distintas longitudes de onda ) presentes
en la membrana tilacoidal.
Una vez captada la energía luminosa por el
complejo antena, esta va a ser transferida a una molécula de clorofila
especial, llamada centro de reacción,
que va a ser la única molécula capaz de ceder un electrón, es decir, de
convertir la energía luminosa en energía química. Así pues, cuando la energía
de la luz llega al centro de reacción, determinados electrones de la clorofila
del centro de reacción son “impulsados” a un nivel energético superior (“excitación
del centro activo”), lo que convierte a esta molécula en un reductor
muy potente. Es así como llegamos al término de Fotosistema (PS), entendiendo por tal al conjunto formado por la
molécula de clorofila del centro de reacción y unas moléculas aceptoras.
Existen dos tipos de fotosistemas:
ü PS I: cuya clorofila se conoce como P700 (por
presentar un máximo de absorción de luz de 700 nm)
ü PS II: cuya clorofila se conoce como P680 (por
presentar un máximo de absorción de luz a 680 nm).
o
Transporte electrónico: dependiendo del recorrido de los electrones, en la
fase luminosa se pueden diferenciar dos tipos de transporte electrónico:
ü Transporte no cíclico (Esquema en Z): en este tipo de transporte los electrones van a
realizar un recorrido abierto, desde el H2O hasta el NADP+,
pasando por el PS II, por una cadena transportadora donde los electrones
“descienden” desde un nivel excitado hasta su nivel normal y finalmente por el
PS I. Se obtiene poder reductor en forma de NADPH.
El mecanismo es el siguiente: tras incidir la luz
sobre el PS I la clorofila del P700 cede un electrón al NADP+ que se
reduce hasta NADPH. La clorofila queda oxidada y debe recuperar el electrón
para volver a ser funcional. El electrón lo recibe como resultado de la
iluminación del PS II que provoca la excitación y emisión de electrones que
viajan por la cadena transportadora hasta la clorofila del PS I. Pero claro,
ahora queda la clorofila del PS II oxidada y debe, por tanto, volver a reducirse;
ello se produce por la rotura (fotólisis)
de una molécula de agua que origina: la cesión de electrones al PS II, la
liberación de H+ al espacio intratilacoidal y la liberación de O2
a la atmósfera.
El flujo que se produce en la cadena de transporte
que conecta los dos fotosistemas provoca la aparición de un gradiente quimiosmótico de H+
entre ambas caras de la membrana tilacoidal, que se empleará para la aparición
de ATP.

ü Transporte
cíclico: (VER ANTES EL SIGUIENTE APARTADO) es
una vía alternativa de la fase luminosa que se produce cuando en los
cloroplastos escasea el ATP y también es típica de bacterias
fotosintéticas anoxigénicas. En ella, un
electrón del PS I es activado por la luz pero, en lugar de viajar hasta el NADP+,
vuelve nuevamente hasta el PS I. En su recorrido es cedido al complejo
citocromo bf, que transporta H+ desde el estroma hasta el espacio
intratilacoidal. Sus características son:
§
Sólo participa el PS I
§
No se produce
reducción del NADP+
§
No hay fotólisis del
agua ni desprendimiento de O2 a la atmósfera.
§
Se produce síntesis de
ATP gracias a la translocación de H+ por el complejo cit bf.
o
FOTOFOSFORILACIÓN: se denomina así al proceso de síntesis de ATP que
se produce, en último término, gracias a la energía contenida en los fotones de
luz. Como ya se comentó antes, el flujo de electrones a favor de gradiente a lo
largo de la cadena transportadora libera energía, que es utilizada para bombear
H+ desde el estroma hasta el espacio intratilacoidal, lo que origina
un gradiente electroquímico de H+ (incrementado por el hecho de que la fotolisis
ocurre en el espacio intratilacoidal, donde se acumulan también los protones
desprendidos) Este gradiente
electroquímico hace que los H+ tiendan a regresar hacia el estroma
y, dado que la membrana tilacoidal es prácticamente impermeable a los H+,
estos solo pueden hacerlo a través de la ATPasa, la cual acopla la energía liberada por el
paso de H+ a favor de gradiente a la fosforilación de ADP, con la
consiguiente síntesis de ATP.

·
FASE OSCURA:
no depende de la luz, ocurre en el estroma del cloroplasto y
consiste en la biosíntesis de compuestos
orgánicos (principalmente glucosa) a partir de CO2, utilizando
la energía (ATP y NADPH) producida en la fase luminosa.
Los bioelementos esenciales para la síntesis de
los monómeros de la materia orgánica (C,N,S y P) se
encuentran en la naturaleza como materia
inorgánica, muy oxidados (CO2, NO3-, SO4=PO4,3-) y requieren un proceso de reducción, con NADPH +
H+ y enlazarlos con ATP(los productos de la fase luminosa) hasta convertirse en
monómeros.

Se
produce mediante una ruta metabólica cíclica llamada “Ciclo de Calvin”. El ciclo consta de tres fases:
·
FASE CARBOXILATIVA: FIJACIÓN DE CO2.
Consiste en la incorporación del átomo de carbono
del CO2 a la ribulosa-1,5-difosfato
(1C+5C), en
una reacción catalizada por la enzima RUBISCO
(ribulosa-1,5-difosfato carboxilasa), la enzima más abundante de la biosfera,
obteniéndose 2 moléculas de 3-fosfoglicerato
(2X3C).
·
FASE REDUCTIVA
Consiste en la reducción del átomo de C que previamente
se ha incorporado y se realiza en dos fases: una fosforilación, del 3-fosfoglicerato hasta 1,3-difosfoglicerato a partir de la
hidrólisis de 1 ATP, y una reducción propiamente dicha, del
1,3-difosfoglicerato hasta gliceraldehído-3-fosfato
a partir de la oxidación de 1 NADPH. Este gliceraldehído-3-fosfato constituye
una encrucijada metabólica, ya que puede seguir distintas rutas:
ü Gluconeogénesis
(una ruta inversa a la glucolisis que permite la obtención de glucosa).
ü Glucolisis, con
la consiguiente obtención de energía.
ü Transformación a piruvato (mediante parte de la glucolisis) que es un
precursor de la síntesis de aminoácidos, o se transforma en Acetil-CoA,
que se utiliza para la síntesis de ácidos grasos.
ü Regeneración de la ribulosa-1,5-difosfato
·
FASE REGENERATIVA
En esta fase el gliceraldehído-3-fosfato se
transforma en ribosa-5-fosfato mediante una serie de reorganizaciones de los
átomos de carbono que originan intermediarios de 3-7 átomos de carbono,
obteniéndose finalmente ribulosa-5-fosfato que, por fosforilación con ATP, se
convierte en ribulosa-1,5-difosfato.
Balance global del Ciclo de Calvin
En cada vuelta de ciclo se reduce
una sola molécula de CO2, por lo que para obtener una molécula de
gliceraldehído-3-fosfato hacen falta tres vueltas; es decir, la reducción de 3
CO2 con el consumo de de 6 NADPH y la energía de 9 ATP. Es decir:
De igual manera, para sintetizar una
glucosa (6C)
se requiere la formación de 2 G3P, por lo que la ecuación global sería: (la
anterior multiplicada por 2)
6 CO2 + 12 NADPH + 12 H+ +
18 ATP → C6H12O6
+ 12 NADP+ + 18 ADP + 18 Pi
FACTORES QUE INFLUYEN EN LA FOTOSÍNTESIS
Son cinco los factores influyentes
en el rendimiento fotosintético:
·
La intensidad
luminosa: por lo general, al aumentar la
intensidad luminosa, aumenta la actividad fotosintética; pero cada especie está
adaptada a unas condiciones óptimas de iluminación (hay plantas adaptadas a
zonas de penumbra y otras a zonas de luz intensa) y superados ciertos límites
se pueden deteriorar los pigmentos fotosintéticos.
·
La concentración
de CO2: la actividad
fotosintética aumenta con la concentración de CO2, hasta llegar a un
máximo en el que se estabiliza debido a la saturación de la enzima Rubisco.
·
La concentración
de O2: al aumentar la
concentración de O2 baja el rendimiento de la fotosíntesis, debido
al proceso de la fotorrespiración.
·
La Tª: la
fotosíntesis se acelera al aumentar la
Tª, hasta un valor determinado, ya que a partir de ese valor
se desnaturalizan las enzimas. Cada especie tiene una Tª óptima en la que la
eficacia de la fotosíntesis es máxima.
·
La humedad: si el aire tiene poca humedad, las plantas
cierran los estomas para evitar pérdidas de agua y ello dificulta la captación
de CO2.
EL PROBLEMA DE LA FOTORRESPIRACIÓN
La enzima Rubisco, además de
funcionar como carboxilasa, puede actuar como oxigenasa (incorporando O2
a la ribulosa-1,5-difosfato en lugar de CO2), siendo esta
oxigenación de la ribulosa-difosfato lo que recibe el nombre de fotorrespiración. El que la enzima actúe
como carboxilasa u oxigenasa depende de las concentraciones relativas de CO2
y O2.
Se le denomina fotorrespiración
porque es dependiente de la luz pero se parece a una respiración, ya que
consume O2 y libera CO2. Tiene lugar cuando la
concentración de CO2 es baja o bien la de O2 alta y
supone una limitación a la eficacia fotosintética dado que al realizarla
la planta consume materia orgánica sin producción de ATP. Debido a ello algunos
tipos de plantas han desarrollado mecanismos para minimizar el problema de la
fotorrespiración:
·
PLANTAS C4 y las PLANTAS
CAM (plantas suculentas = acumulan agua en tallos y hojas): se trata de plantas típicamente de climas
cálidos, que durante el día cierran sus estomas para evitar la deshidratación.
Pero claro, esto también conlleva una disminución de la concentración de CO2 y un aumento de O2 ya que la
fotosíntesis continúa. Por tanto, lo que hacen es que durante la noche abren lo
estomas (evitando la pérdida de agua) y absorben CO2 que almacenan
en forma de ácido málico (4C)
que se incorpora al Ciclo de Calvin durante el día.
La diferencia entre ambas plantas es que en las C4
la fijación del CO2 en ácido málico y el Ciclo de Calvin están
separados espacio-temporalmente (estas plantas presentan una anatomía foliar
diferente: en las células del mesófilo ocurre la fijación del CO2 y
el Ciclo de Calvin en las células de la vaina) mientras que en las plantas CAM sólo hay separación
temporal.
No hay comentarios:
Publicar un comentario